

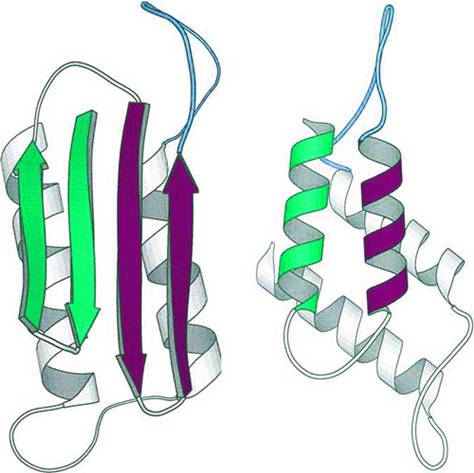
Les formes saines et pathologiques du prion diffèrent par la configuration de certaines parties
L’immense complexité du vivant a pu faire croire que les méthodes de la physique ne pouvaient être utiles pour le comprendre. Avec le recul, il est apparu que cette difficulté était due au fait que les systèmes biologiques évoluent dans un univers impliquant un très grand nombre d’états d’énergies voisines et séparés par des barrières peu élevées : une faible contrainte imposée peut entrainer un déplacement considérables des équilibres.
Un problème typique est le repliement des protéines. Le paradoxe de Levinthal résulte simplement du dénombrement des configurations géométriques possibles d’une protéine moyenne, de l’ordre de 10 ^ 143 ! Il est donc impossible qu’elle explore toutes les configurations pour trouver la plus stable. Il est également possible qu’il existe plusieurs états métastables. In vivo, une mauvaise configuration de certaines protéines est considérée comme la cause de pathologies graves (ESB, Creutzfeld-Jacobs, Alzheimer).
Nous avons étudié une facette de ce problème en considérant un système "modèle" i.e. considérablement simplifié : un peptide composé de la répétition d’un seul acide aminé, l’acide polyglutamique. Cet homopeptide est connu pour sa propension à constituer une hélice à basse température et une chaîne aléatoire à température élevée. L’état hélicoïdal peut être facilement quantifié expérimentalement par la technique de dichroïsme circulaire. Cette transition a été modélisée dès 1959 par une théorie mettant en jeu deux paramètres énergétiques, l’énergie d’une boucle élémentaire et l’énergie de couplage entre boucles voisines ainsi que l’entropie des configurations. Ce modèle est formellement analogue au modèle d’Ising à une dimension et donc soluble. Des mesures récentes précises se sont révélées incompatibles avec ce modèle. Nous avons pu montrer que par contre les données étaient bien décrites par la considération d’un troisième état possible et l’absence de coopérativité.
Un autre aspect du problème est d’essayer de guider le repliement d’une protéine complexe (qui ne se replie pas spontanément dans sa configuration fonctionnelle) en s’inspirant du vivant. Les protéines synthétisées séquentiellement acquièrent leur structure également séquentiellement après être passé dépliées au travers d’un pore moléculaire nanométrique (translocation). Pour simuler ce procédé, nous avons tenté d’utiliser des pores nanométriques obtenus par la méthode de "track-etch" dans des membranes plastiques, sans résultat, ce que nous attribuons à l’interaction des protéines avec les charges électriques liées laissées à la surface du pore après l’attaque chimique. Nous entreprenons donc maintenant l’élaboration de membranes métalliques présentant des pores comparables en utilisant les membranes actuelles comme gabarit.
